The subspecies was described and named by Albert Schwartz (1923 – 1992). He was a Professor of Biology at Miami-Dade Community College. Schwartz co-authored the seminal work on the West Indian boas "Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships."
Holotype
CM no. 60519, adult female taken on 10 August, 1976 by David A. Daniels. Paratype ASFS V10312, taken by R. Thomas on 31 March, 1966. Same locality as the holotype.
The name manototus is derived from the Greek for "most rare" .
Common Name
Île à Cabrit Boa.
Description and taxonomic notes
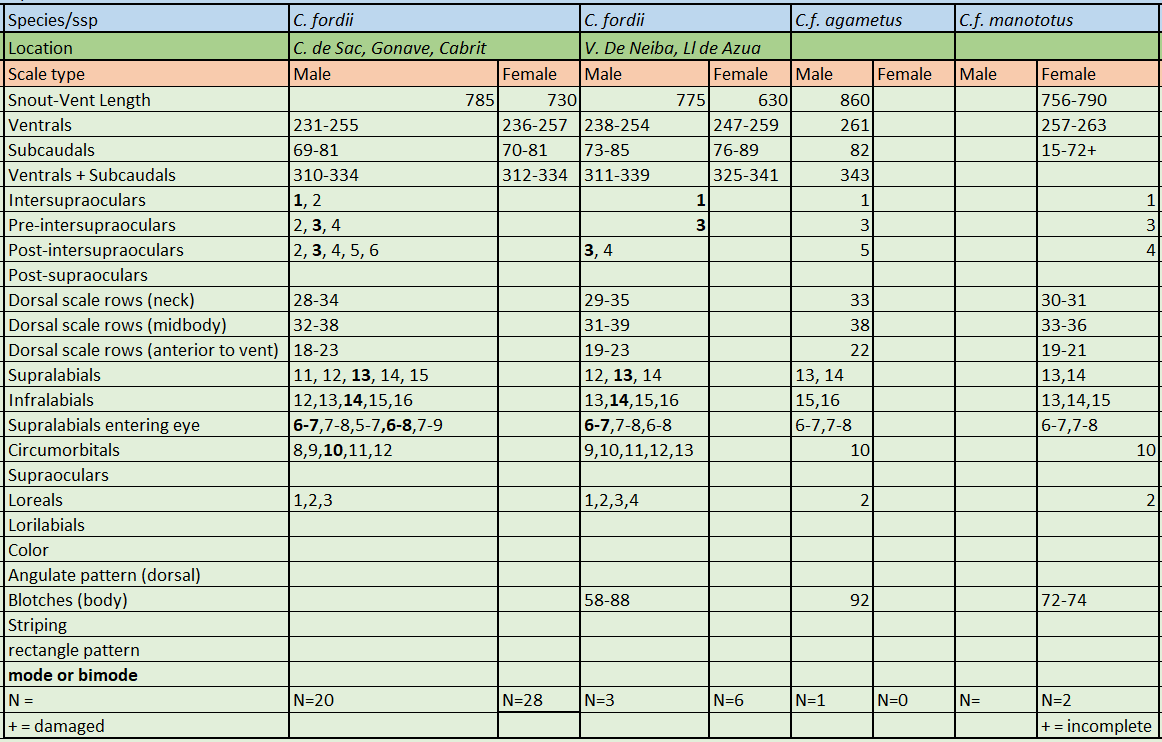
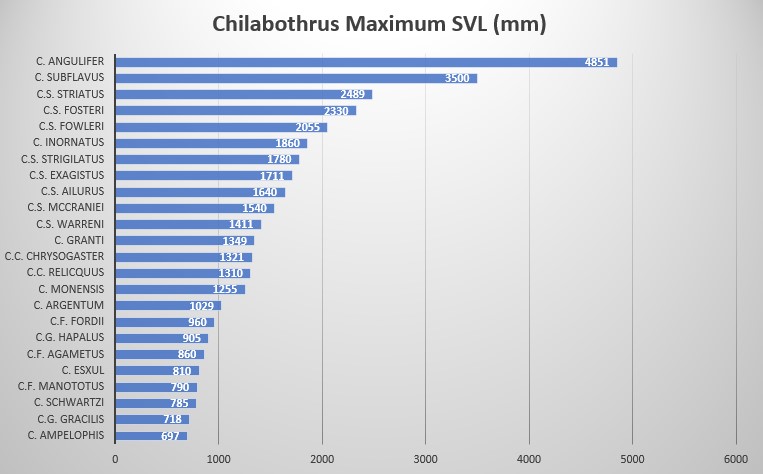
Chilabothrus f. manototus is sexually dimorphic. In females it is characterized by a SVL of 790 mm, ventral scales are 263, subcaudal scales 29 (incomplete), scale rows at neck are 30, scale rows at midbody are 36, scale rows anterior to tail are 21, supralabials are 14-13, infralabials are 13-14, circumorbital scales are 10, supralabials entering the eye are 6-7 on one side, 7-8 on the other side, loreals are 2-2, preintersupraoculars are 3, intersupraocular is 1, postintersupraoculars are 4. Dark gray dorsal ground color lightening on the lower sides to pale gray. 72-74 dorsal blotches red-brown with brown edges; each blotch surrounded by pale gray . C.f. manototus and C.f. agametus are easily differentiated by the number of dorsal blotches; 72-74 in C.f. manototus and 92 in C.f. agametus. Chilabothrus fordii is one of the smallest species within the genus. Neonate C.fordii (and C. monensis) are the only members of the genus born with a gray or grayish-brown ground color .
The two subspecies Chilabothrus fordii agametus and C. f. manototus were described by Sheplan and Schwartz (1974) and Schwartz (1979) on the basis of one C. f. agametus and two C. f. manototus specimens. While a small sample size should in general not hinder the description of a species or subspecies, it is crucial to remark at this point, that some doubt lasts over the justification of both subspecies.
Reasons are: In case of C. f. manototus the phollidosis is within the phollidic range of the nominate subspecies. The islet Ile a cabrit is very small (.25km2) and is located only about 200m off the coastline of Hispaniola.
The fact that only two specimens were found and that inhabitants of the island reported to have never seen such animals at all, opens the possibility for an alternative explanation. The species is not naturally occurring at the islet and that the two specimens found came to Ile a Cabrit through different sources, potentially from the Dominican Republic with plants destined for the beach resort Ibo Beach Club, which opened in 1964 and changed the island flora massively. Should this last scenario be true, the possibility that C. f. manototus is a valid subspecies is still possible, yet the type locality would be different.
However, the current situation makes it hard to justify the status of the subspecies agametus and manototus. We consider it necessary to analyse morphologically as well as genetically C. fordii from different localities across the range to clarify the subspecific question.
Distribution
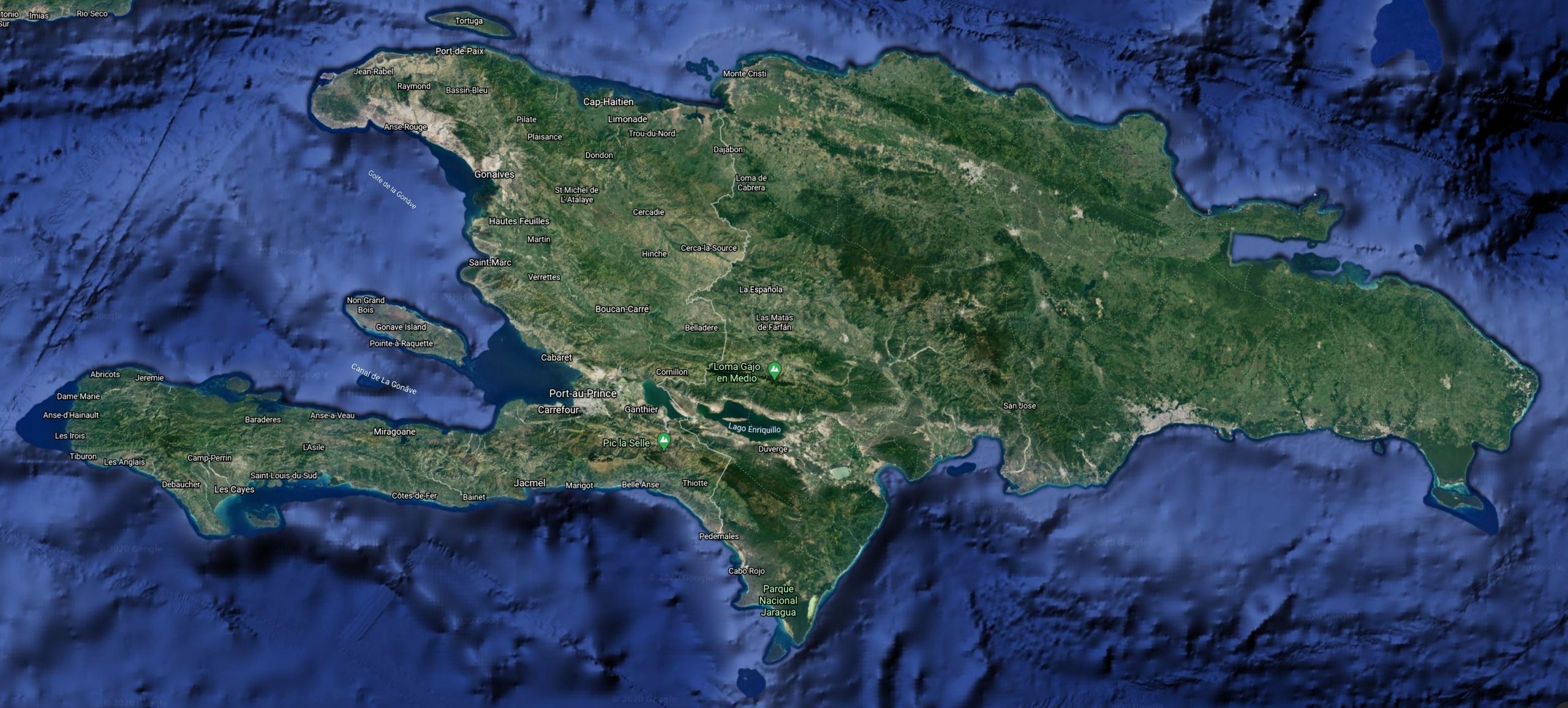
Known only from Île à Cabrit, Departement de l’Ouest, Haiti.
Three subspecies of Chilabothrus fordii occur on Hispaniola. The Flags on the map indicate a type locality of each subspecies. Scroll over to see which subspecies occurs at the indicated point.
Habitat
Ile a Cabrit appears to be a microenvironment on an island off the coast Hispaniola only 0.25 km2 in size and 500m away from the coast of Hispaniola. Schwartz noted that the flora of the island has massively changed since the opening of the popular Ibo Beach Hotel in 1964. He noted that there still remain some vestiges of the original vegetation primarily in areas remote from the hotel. Schwartz visited the island twice, even though both visits were only for portions of a day, he noticed that at the first visit the island was dry and dusty, vegetation was brown, and the general aspect was forbidding. During his second visit, in 1977, conditions were more mesic and the islet seemed less rigorous. During both visits he was impressed with the high temperatures and high humidity.
The Island (especially the vegetation and the climeate ) might have changed since Schwartz visited it in 1977. He wrote . :
Ile a Cabrit is roughly circular with a western peninsula composed of mangroves and without apparent dry land. The island has a hillock on the eastern end that rises to about 25 m. The periphery is mostly mangrove-fringed and backed in places along the northern coast by some xeric deciduous and thorn forest. There is also a patch of apparently original woods in the area of the swimming pools east of the hotel near the southern coast. Most of the islet apparently was once covered with grasses and grassy tussocks. Much of the latter have been cut and the tussocks and cut grass have been left in numerous piles from the hillslope to near the shore. There are limited stands of grasses still remaining especially behind the north coast woods. The island is barren with a minimum of shade and there are few places for reptiles to find cover except for the cut grasses, an occasional dry and decayed tree or arborescent cactus trunk, and some debris from the hotel (e.g., decayed piles of boards used for concrete construction forms). Ile a Cabrit thus does not appear encouraging to someone interested in collecting its herpetofauna. I assume that originally Ile a Cabrit supported a xeric flora with some peripheral shaded deciduous woodlands and a mangrove border. The area about the hotel is landscaped and moderately shaded and watered, but in dry 1976 it still had a barren and desolate look.
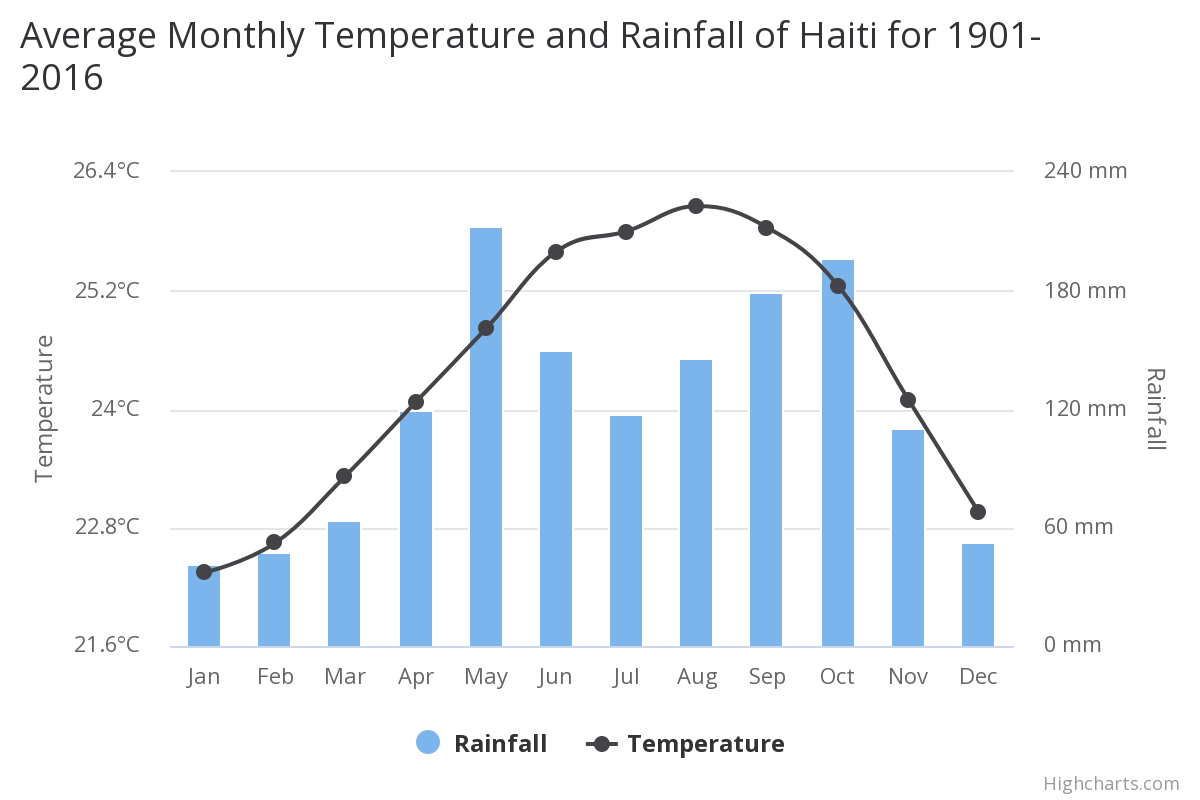
We have no weather or temperature data for Ile a Cabrit, however overall on Haiti mean temperatures have increased by 0.45°C since 1960, with warming most rapid in the warmest season, June-November. The frequency of hot days and hot nights increased by 63 and 48 days per year, respectively, between 1960 and 2003. The frequency of cold days and cold nights has decreased steadily since 1960. Mean annual rainfall has decreased by 5 mm per month per decade since 1960. The intensity of Atlantic hurricanes has increased substantially since 1980.
Longevity
No age records for C. f. manototus have been reproted. We assume that the subspecies is long lived, like C. f. fordii.
Reproduction
No data regarding the reproductive biology of this subspecies exists. We assume this taxon has a similar reproductive biology as the nominate subspecies. Based on the weather, geography and proximity to the nominate species, it would not be unreasonable to attribute the same mating behavior, gestation and parturition data to the subspecies. Locality differences in climate and habitat may influence the start and end period for mating and parturition.
Behavior
Nothing regarding the behavior of C. f. manototus has been published. In regards to the xeric nature of the island and the limited vegetation we assume that the behavior is similar to the nominate subspecies
Diet
Nothin has been published about the diet of C. f. manototus. We assume that the diet is similar to that of C. f. fordii on the mainland where the nominate subspecies preys on Anolis and small rodents (Mus) . An examination of the stomach contents of 48 C. fordii revealed that boa preys upon Anolis cybotes, Anolis sp. and small rodents (Mus) . On Ile a cabrit 10 lizard species occur (Anolis brevirostris, Anolis olssoni, Anolis semilineatus, Anolis cybotes, Ameiva lineolata, Ameiva chrysolaema, Leiocephalus pratensis, Aristelliger cochranae, Sphaerodactylus altavelensis, Celestus stenurus) and the only snake species that has been found is C. f. manototus . The lizard species possibly function as prey for the boa. It is unclear if Mus musculus occurs on the island, given that Schwartz described the environment as very dry and xeric with higher temperatures than on the main Island Hispaniola, it is possible that mice can’t survive under these conditions. In this case, the boas would thrive entirely on a saurophagous (or avian?) diet.
Captive management and captive population
To our knowledge, the Ile a Cabrit Boa has not been kept in captivity. We assume the same parameters as for the nominate subspecies would benefit these boas. Possibly adjusting the climate regimen to the environment from Ile a Cabrit.
Conservation status, threats and population size in nature
CITES: Appendix II
Haiti is not a member of CITES.
Dominican Republic joined CITES on 17 December, 1986; entry into force on 17 March, 1987.
We have no information about the current situation on Ile a Cabrit. Judging from the Satellite images (see below) we assume that the island is still covered by some vegetation, however on the southern side of the island, the landscape seems largely degraded. C. fordii has previosly been found to be sustaining viable populations, sometimes even in heavily disturbed areas . This give some hope for the presence of C. f. manototus on the island.
In general the threats on Hispaniola are plentiful and reach from introduced Bufo marinus, a species that has become widely distributed and is invading even very xeric areas , to habitat destruction through deforestation and soil erosion. Overpopulation leads to inadequate supplies of potable water and and a lack of sanitation; natural disasters have been on the increase .
The 2010 earthquake left Haiti largely ungoverned and the humanitarian crisis was followed by an ecological crisis due to destruction of habitats. Thus making it difficult to envisage a stable future for the snake species on the island. In addition to this, man made climate change is a factor which is likely to gain momentum in ecosystem changes throughout the West Indies. Giant storms fueled by rising oceanic temperatures as a consequence of human caused climate change are more likely to form more often than before. On the other hand, climate change will also increase droughts and diminish fresh water resources on the islands of the West Indies.
In a relatively recent study by Hedges et al. they predict, through modeling, that Haiti will experience mass extinction events by 2036 as a result of current deforestation trajectories. They comment: “We project that all primary forest in Haiti will disappear by ∼2035 (CI, 2033.5–2035.4) at the current rate, defining phase II as a 49-year period from 1986 to 2035 during which the final 4.8% of primary forest in the country will be lost. Assuming that our estimated loss of vertebrates is representative of the biodiversity in general, then 66–83% of species will be lost in Haiti during 1986–2035 because of deforestation. Thus, Haiti is well into a mass extinction, with only 8 of 50 mountains still holding primary forest. Unfortunately, Haiti’s neighbor, the Dominican Republic, is not a major refuge for Haitian species because more than half of the species surveyed (51%, average) on mountains with primary forest are endemic to Haiti and 12% are endemic to an individual mountain in Haiti. In addition, forest loss in the Dominican Republic is a threat to that country’s biodiversity.”
They continue: “It is common for deforestation to occur within protected areas of tropical countries, and, during the course of our surveys in Haiti, we observed ongoing destruction of primary forest inside all of the national parks. We also estimate that 60–75% of primary forest in the two original national parks, Pic Macaya and La Visite, has disappeared since they were declared as protected areas 35 years ago. In both cases, the rates of primary forest loss (pre-2000 and post-2000) were greater than the overall rates for all of Haiti, indicating that protection was minimal or nonexistent. This indicates that the mass extinction of biodiversity in Haiti will continue unabated unless greater protective measures are taken. More generally, this suggests that the phrase “protected area” be reserved only for areas where protection has been confirmed.” , .
A study on the risks for species extinction on island ecosystems worldwide analysed 15 insular regions, applying novel network analysis methods which enabled to disentangle the associations of multiple threats on vertebrates, invertebrates, and plants. The results showed that Biological invasions, wildlife exploitation, and cultivation, either alone or in association, are the three most important drivers of species extinction and decline on islands to date . These results are consistent with expectations when analysing past and even current events. However, considering that climate change will dramatically change whole ecosystems in previously unprecedented ways, we estimate that this threat is more likely to affect species survival on an overall scale than the past drivers of extinction.
For a nation struggling to feed and house its people, conserving its fauna and herpetofauna are far from the top of the list, if they are on the list at all. Granted, there are measures in place, all on paper, that lay land aside for National Parks. Yet enforcing the measurements laid out on paper are anything but enforceable, as noted above. It is truly a travesty of the highest order; we can only hope some of the herpetofauna escapes the sickle of extinction.
Thus we consider the biggest threads for species survival in (1) large scale ecosystem changes, (2) invasive species, (3) climate change.
We feel, given the boas’ extremely small range and the current rate of habitat destruction taking place it is imperative to determine the current status of the three subspecies as quickly as time and resources allow.
The maps below illustrate the extent of habitat destruction and alteration due to development, intentional deforestation and agriculture.
Ile a Cabrit, HaitiHispaniola (C.f. fordii, C.f. agametus, C.f. manototus, C.g.gracilis, C.g. hapalus, C. s. striatus, C.s. warreni, C.s. exagistus)Early map of Hispaniola, 1725.
On display in these Zoos
We are unaware of any live Chilabothrus fordiimanotous in Zoo or Academic collections anywhere in the world. If you have further information, please Contact us.
Hedges, S. B., Cohen, W. B., Timyan, J., & Yang, Z. (2018). Haiti’s biodiversity threatened by nearly complete loss of primary forest. Proceedings of the National Academy of Sciences, 201809753. https://doi.org/10.1073/pnas.1809753115
Hedges, S. B., Powell, R., Henderson, R. W., Hanson, S., & Murphy, J. C. (2019). Definition of the Caribbean Islands biogeographic region, with checklist and recommendations for standardized common names of amphibians and reptiles. Caribbean Herpetology, 1–53. https://doi.org/10.31611/ch.67
Henderson, R. W., & Powell, R. (2004). Epicrates fordii. In Catalogue of American Amphibians and Reptiles (CAAR).
Henderson, R. W., & Schwartz, A. (1984). A guide to the identification of the amphibians and reptiles of Hispaniola (Vol. 4). Milwaukee Public Museum; /z-wcorg/.
Henderson, R., Noeske-Hallin, T. A., Ottenwalder, J. A., & Schwartz, A. (1987). On the Diet of the Boa Epicrates striatus on Hispaniola, with Notes on E. fordi and E. gracilis. Amphibia-Reptilia, 8, 251–258. https://doi.org/10.1163/156853887X00298
Leclerc, C., Courchamp, F., & Bellard, C. (2018). Insular threat associations within taxa worldwide. Scientific Reports, 8(1). https://doi.org/10.1038/s41598-018-24733-0
Powell, R., & Incháustegui, S. (2009). Conservation of the herpetofauna of the Dominican Republic. Applied Herpetology, 6(2), 103–122. https://doi.org/10.1163/157075409X427153
Powell, R., Ottenwalder, J. A., Inchaustegui, S. J., Henderson, R. W., & Glor, R. E. (2000). Amphibians and reptiles of the Dominican Republic: species of special concern. Oryx, 34(2), 118–128. https://doi.org/10.1046/j.1365-3008.2000.00103.x
Reynolds, R. G., & Henderson, R. W. (2018). Boas of the World (Superfamily Booidae): A Checklist With Systematic, Taxonomic, and Conservation Assessments. Bulletin of the Museum of Comparative Zoology, 162(1), 1–58. https://doi.org/10.3099/MCZ48.1
Schwartz, A. (1979). The Herpetofauna of Ile à Cabrit, Haiti, with the Description of Two New Subspecies. Herpetologica, 35(3), 248–255. http://www.jstor.org/stable/3891695
Schwartz, A., & Henderson, R. W. (1991). Amphibians and Reptiles of the West Indies: Descriptions, Distributions, and Natural History. University of Florida Press.
Sheplan, B. R., & Schwartz, A. (1974). Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships. Annals of the Carnegie Museum, 45, 57–143.