Holotype of Chilabothrus striatus warreni. Photo by MCZ
The subspecies was described and named by Sheplan & Schwartz. Bruce R. Sheplan was a herpetologist at Miami-Dade Community College. Albert Schwartz (1923 – 1992) was a Professor of Biology at Miami-Dade Community College. Both co-authored the seminal work "Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships."
Holotype
MCZ no. 125604 (originally ASFS V20364) from vicinity Ile de la Tortue (Palmiste), Haiti. Captured by a local collector for C.R. Warren on 15-27 August, 1970. Paratypes ASFS V20367-78, young removed from MCZ 125604; USNM 59437 Ile de la Tortue on 1 February, 1917 by W.L. Abbott.
The name striatus is derived from the Latin for "striated" or "lined."
Sheplan & Schwartz named C.s. warreni after C. Rhea Warren, who collected extensively on Ile de la Tortue, Haiti.
Common Name
Chilabothrus striatus warreni: Ile de la Tortue Boa.
Description and taxonomic notes
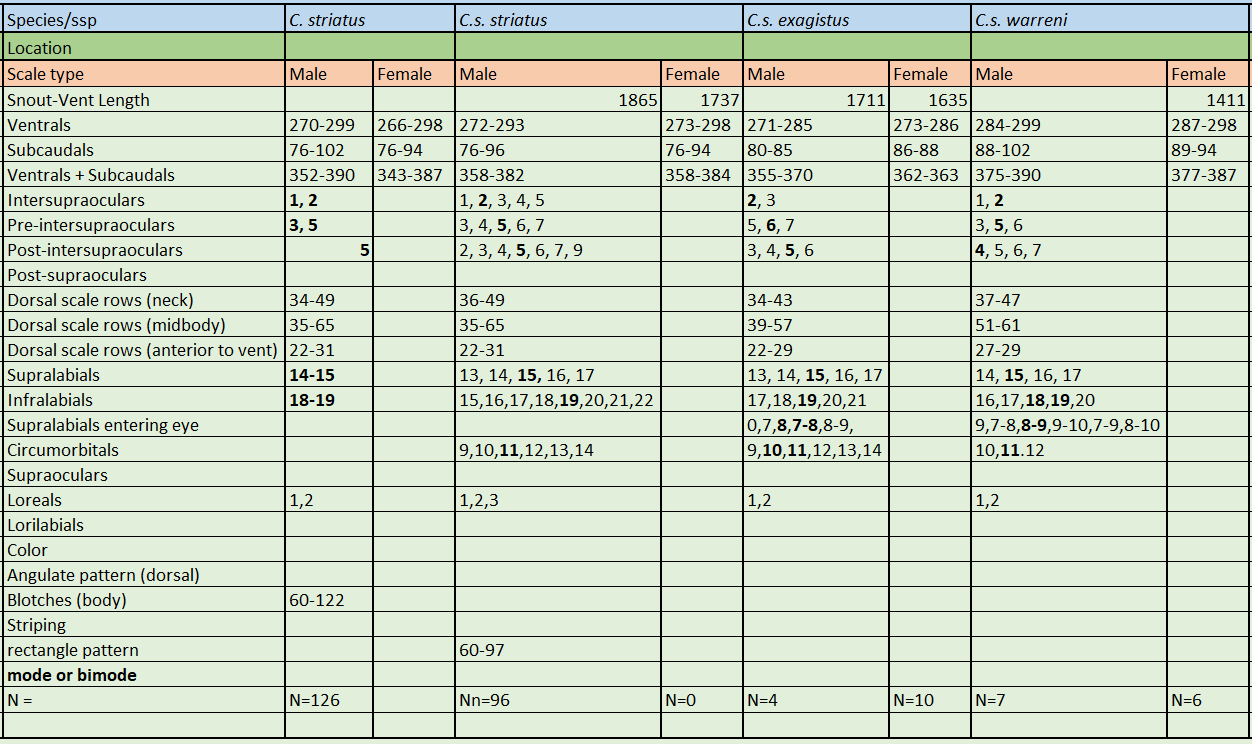
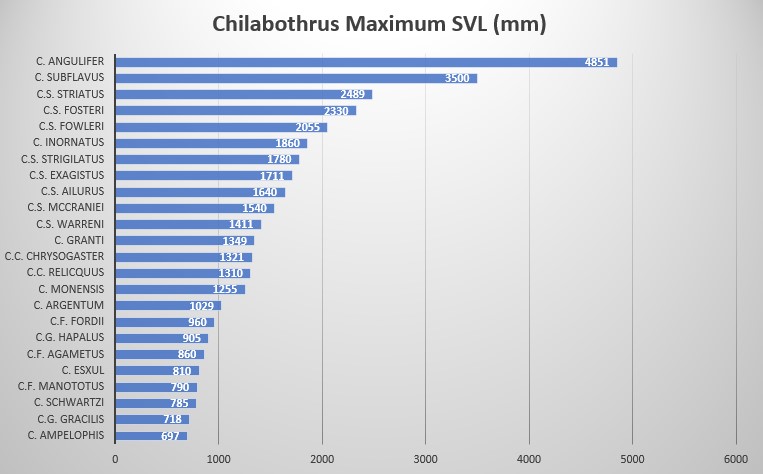
Chilabothrus striatus warreni is presumably a moderately sized boa with the females reaching 1411 mm SVL. There are no adult males in any museums. thus no adult size data. It is characterized by high ventral counts in both sexes; males are 284-299 ventrals and females are 287-298 ventrals. 2 superintraoculars (modally) bordered by 5 scales anteriorly and 4 scales posteriorly (modally), supralabials 15 (modally) with 2 supralabials entering the eye (8-9), 11 scales in the circumorbital series (modally) and 2 loreals (modally).
The specimens from Tortue were examined in life by Warren and were pale dorsally. It lacks the neck stripe, secondary lateral blotching and there is extensive fusion of the anterior pattern. C.s. Warreni differs from C.s striatus and C.s. exagistus in that:
All three subspecies show a differing in dorsal pattern intensity with C.s. striatus the darkest, C.s. exagistus the lightest and C.s. warreni in between the two.
Only C.s. warreni possesses, on a regular basis, median neck spots or dots.
The head scale formula differs for all three subspecies; C.s. warreni is 5-2-4, C.s striatus is 5-2-5 and C.s. exagistus is 6-2-5.
C.s. warreni is separated from C.s. exagistus by the higher number of ventrals + subcaudals.
C.s. warreni and C.s. striatus have 2 loreals vs. one in C.s. exagistus.
C.s. warreni and C.s. striatus possess supralabials involved in eye penetration of 8-9 while C.s. exagistus has 7-8.
Holotype of C. s. warreni. Photo by MCZHolotype of C. s. warreni. Photo by MCZC.s. striatus, C.s. exagistus, C.s. warreni meristics. *
* Source
Cochran investigated a specimen from Isla Tortuga but found that “The specimen from Tortuga Island does not seem to differ specifically from the Haitian form.” About 50 years later Sheplan and Schwartz considered the differences between this population and the mainland form so significant that the erection of the subspecies warreni was reasoned . Another 19 years later Robert Powell was fascinated by Hispaniola and its satellite islands as they are a microcosm of various biomes, often only a few kilometers apart. Applying the criteria of the evolutionary species concept, he found that 43 species on Hispaniola might match the criteria for being species complexes as 1) they are distinctly mapped as allopatric, and 2) they have been formally diagnosed as being morphologically distinct. Thus, he listed recognized Hispaniolan subspecies which may meet criteria for recognition as species by application of the evolutionary species concept. Among these were six snake species, one of which is Chilabothrus striatus warreni . Six years later, Crother supported this view citing Powell . It will be interesting to see if molecular data combined with further morphological studies corroborate this opinion.
A molecular phylogeny based on CYTB locus found that Chilabothrus striatus is most closely related to C. exsul, C. schwartzi and C. argentum: the lineages split about 2.2 Mya. The next closest relative to the clade is C. strigilatus, from which the lineages split 2.6 Mya. Then there is C. chrysogaster, from which the lineages split 4.1 Mya .
Holotype of C. s. warreni. Photo by MCZHolotype of C. s. warreni. Photo by MCZHolotype of C. s. warreni. Photo by MCZHolotype of C. s. warreni. Photo by MCZ
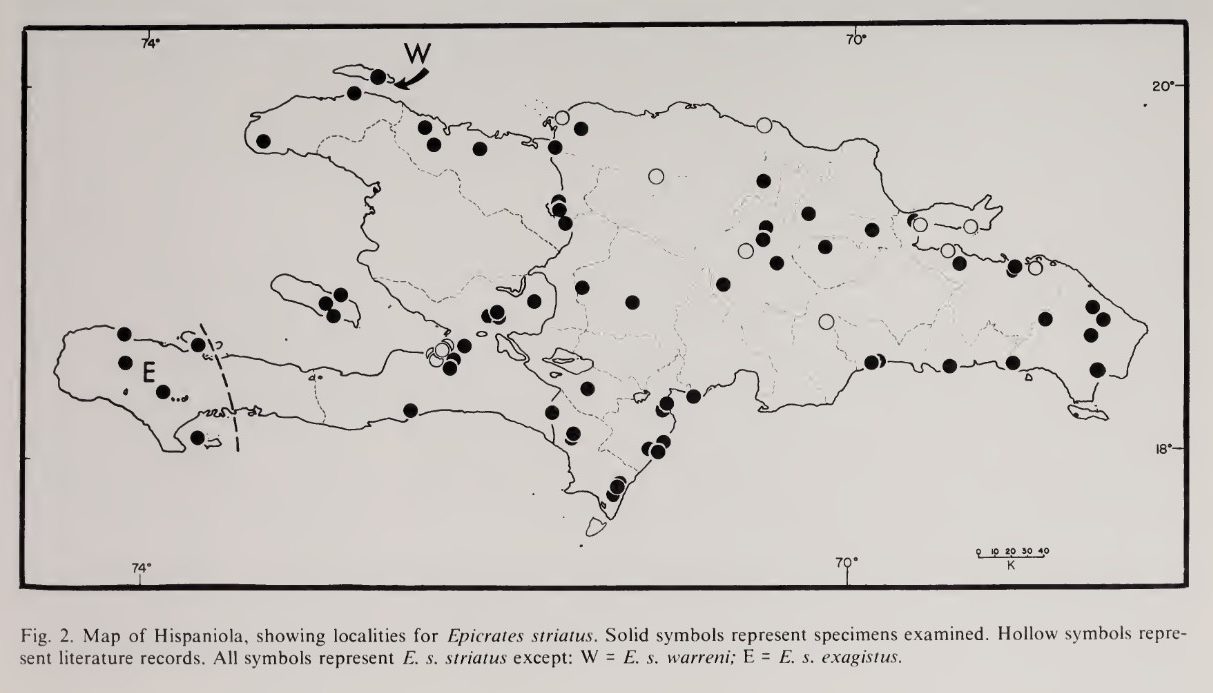
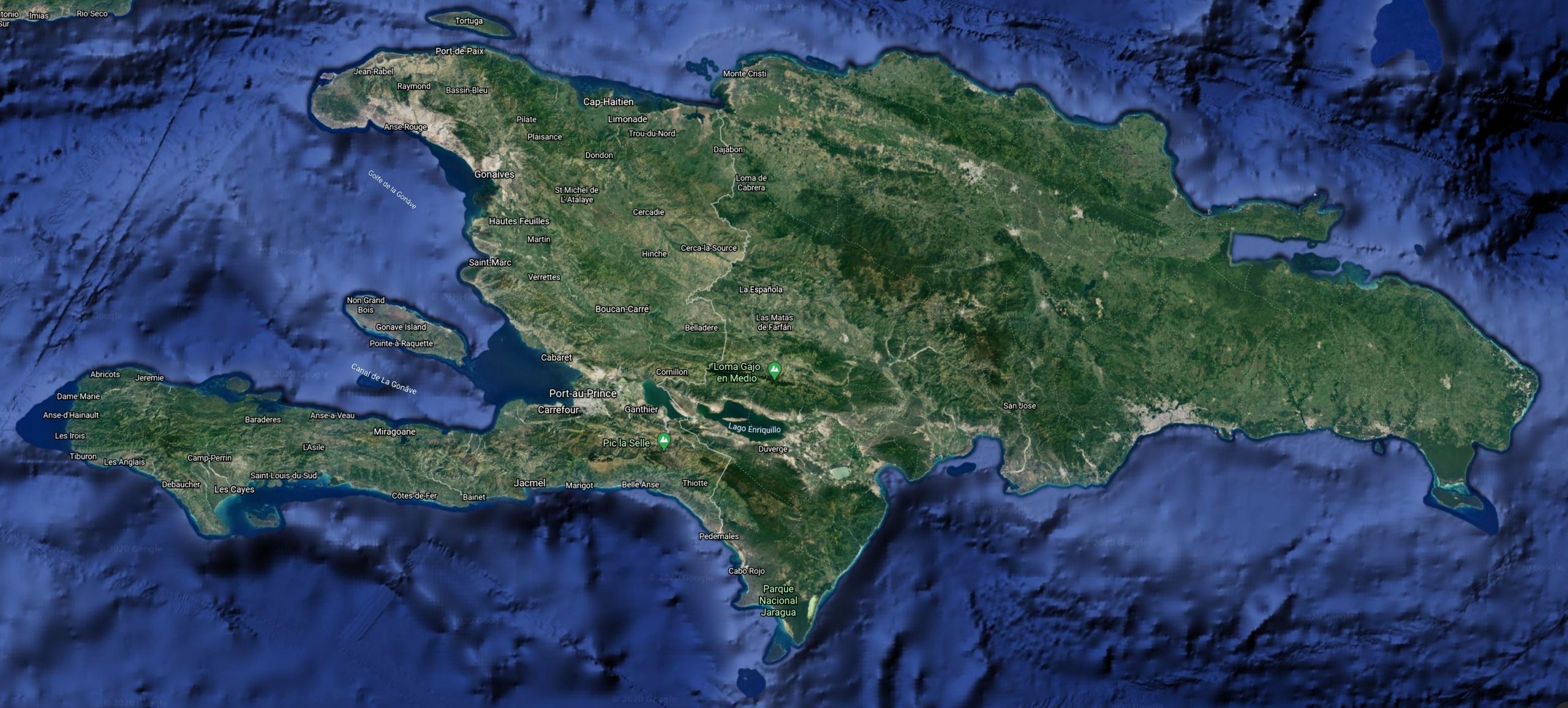
Distribution
C.s. warreni: Haiti, Île de la Tortue.
Distribution map of C.s. striatus, C.s exagistus & C.s. warreni. *
* Source
Three subspecies of Chilabothrus striatus occur on Hispaniola. The flags on the map indicate the type locality of a subspecies. Scroll over to see which subspecies has its type locality at the indicated point.
Habitat
The island of Tortuga stands off the northern coast of Haiti. Overall the island is mountainous; the northern part is very rocky which is why the northern shore was considered as inaccessible via both land and sea. Today small settlements occur all over the island and the total human population is about 26000 (census 2003). Many areas are now under deforestation pressure.
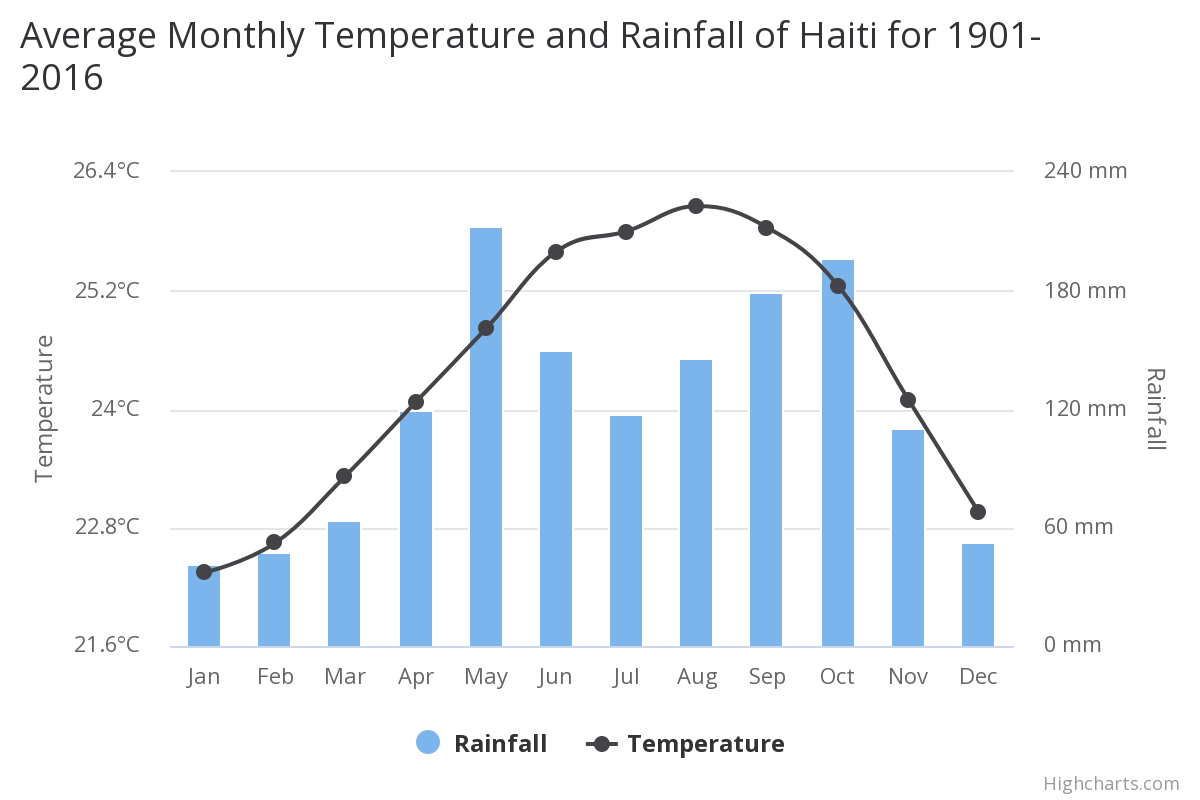
On Haiti the mean annual temperature is 24.40°C and the mean annual precipitation is 1438.40 mm. Located in the Caribbean’s Great Antilles, Haiti has a hot and humid tropical climate. Daily temperatures typically range between 19°C and 28°C in the winter and 23°C to 33°C during the summer months. Northern and windward slopes in the mountainous regions receive up to three times more precipitation than the leeward side. Annual precipitation in the mountains averages 1,200 mm, while the annual precipitation in the lowlands is as low as 550 mm. The Plaine du Gonaïves and the eastern part of the Plaine du Cul-de-Sac are the driest regions in the country. The wet season is long, particularly in the northern and southern regions of the island, with two pronounced peaks occurring between March and November.
Mean temperatures have increased by 0.45°C since 1960, with warming most rapid in the warmest season, June-November. The frequency of hot days and hot nights increased by 63 and 48 days per year, respectively, between 1960 and 2003. The frequency of cold days and cold nights has decreased steadily since 1960. Mean annual rainfall has decreased by 5 mm per month per decade since 1960. The intensity of Atlantic hurricanes has increased substantially since 1980.
C.s. warreni is thought to inhabit the same mesic forests as its congeners. Schwartz quotes Warren in that Palmiste is (or was) a mesic region .
Longevity
C. striatus are long lived. Wagner (pers. comm.) has an old male from one of the authors that was an adult when imported in the early 2000’s. Bowler listed the maximum age for a Epicrates striatus ssp. with 22 years and one month . While we can not exclude with certainty that this record refers to C. striatus, we assume that this record refers to a Chilabothrus strigilatus ssp., since (to the best of our knowledge) no Chilabothrus striatus warreni and very few C. s. exagistus have been exported and kept. The only age record we could find about C. s. striatus confirms an age of 6 years and 8 months at the National Zoological Park .
Reproduction
The only note on reproductive biology stems from the holotype which contained eleven juveniles which were then quite well developed and active despite the presence of large amounts of yolk. One juvenile was deformed. The holoptype was collected in August 15 – 27, 1970 . This suggests that the parturition occurs in nature in beginning – mid September. The number of young from the holotype is in the same range as the nominate species.
Behavior
No field data or captive observations have been made from C. s. warreni. We assume it does not differ significantly from the nominate subspecies, yet, the high ventral count might indicate a more arboreal lifestyle. The presumed occurrence of this boa in mesic forests corroborates this theory. We assume that it is – like the nominate species – nocturnal.
Diet
Henderson et al. examined 133 C. striatus and found 28 prey items. The boas go through an ontogenetic shift in prey items as they mature. Boas that measure less than 60 cm SVL preyed primarily on Anolis. Boas measuring 60 to 80 cm SVL preyed on both Anolis and small rodents. Those boas larger than 80 cm SVL preyed on birds and rats (Rattus rattus). Once boas’ measurements exceeded 105 cm SVL their diet consisted entirely of rats. In 1974 an extremely large C. striatus (248.9 cm total length) was found that contained remains of a Plagiodontia in its digestive tract (C.A. Woods, reported in Ottenwalder, 1985). Bats are preyed upon, but only where they are localized geographically. The prey items of the 133 boas examined consisted of:
2.1% lizards (Anolis coelestinus, A. cybotes, A. sp.)
It was determined the mean size of anoles taken (including C. fordii and C. gracilis), to be 3.6 cm³ (2.5 to 4.6 cm³). C. striatus preyed upon the larger anoles (of a species) more so than any other boid species. They posit the large boas feed only upon large prey because the prey is both ecologically and geographically abundant and widespread .
Captive management
No information regarding the captive management of C. s. warreni are available.
Conservation status, threats and population size in nature
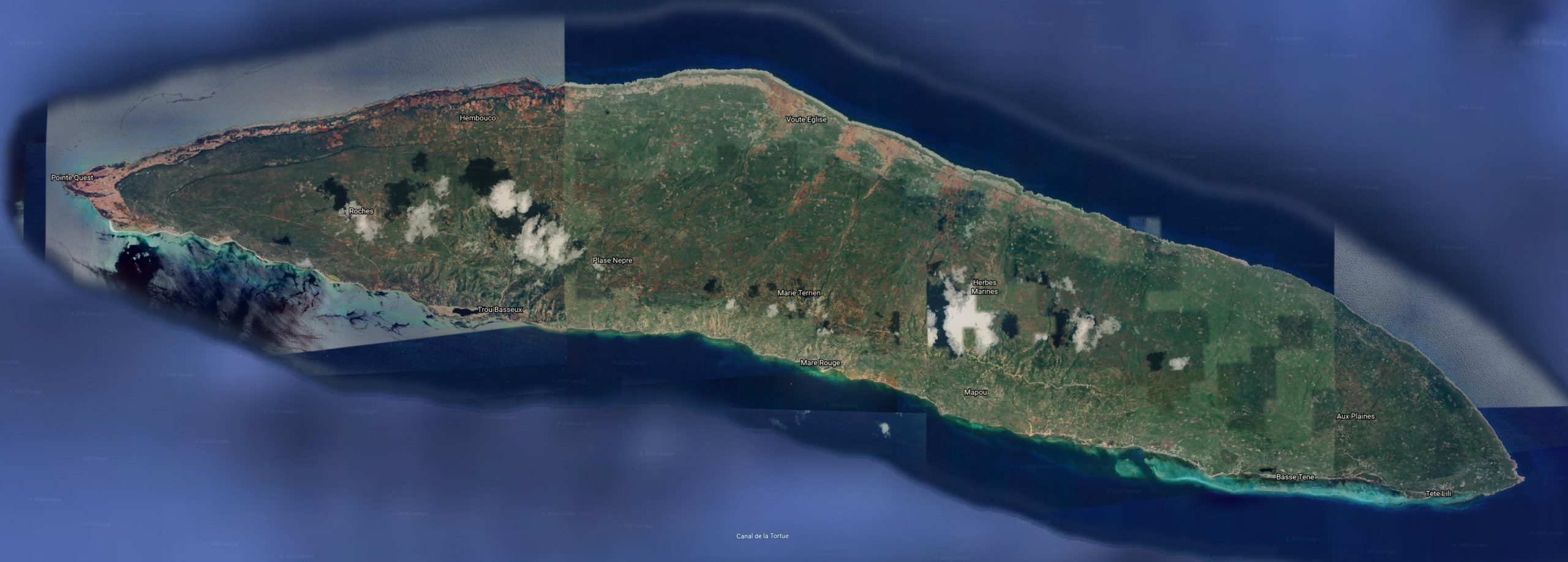
No information regarding the abundance of C. s. warreni are available. Haiti became one of the poorest countries in the world from its once flourishing state and is now suffering from high degrees of poverty. As a result Haiti is largely deforested, since wood and charcoal are the primary source of energy. Analysis of satellite images confirms that deforestation is an issue on Tortuga Island as it is on the rest of Haiti. Patches of bare red soil indicate the deforestation and erosion in place. The conservation efforts undertaken in the previous years in Haiti have largely fallen victim to inactivity by the fractured departments within the unstable government. Travel to and within Haiti is considered high risk. The recent earthquake and hurricane tragedies have compounded all of Haiti’s problems. Habitat destruction will remain an issue and subsequent soil erosion will lead to further problems.
The CIA World Factbook lists the following environmental threats for Haiti: extensive deforestation (much of the remaining forested land is being cleared for agriculture and used as fuel); soil erosion; overpopulation leads to inadequate supplies of potable water and and a lack of sanitation; natural disasters .
The map below illustrates the extent of habitat destruction and alteration due to development and agriculture.
Hispaniola (C.f. fordii, C.f. agametus, C.f. manototus, C.g.gracilis, C.g. hapalus, C. s. striatus, C.s. warreni, C.s. exagistus)Tortuga Island or Ile de la Tortue. (Chilabothrus striatus warreni)
Early map of Hispaniola, 1725.
Population in captivity
We are unaware of any C.s. warreni in any Public Institutions or private collections worldwide.
On display in these Zoos
None that we are aware of.
Anecdotal information
Doris M. Cochran wrote, in 1924, of Dr. W.L. Abbott:
“For the past thirty-five years Dr. W. L. Abbott has enriched the collections in the United States-National Museum by frequent contributions of the results of his collecting expeditions in various parts of the world. Since 1916 he has turned his attention particularly to the island of Haiti, from which he has sent much valuable material,
including many new or rare species of animals and plants. During the summer and autumn of 1916 Doctor Abbott collected natural history specimens on the Samana Peninsula in northeastern Santo Domingo. This trip proved so beneficial to the needs of the National Museum that Doctor Abbott has returned to the island each year. His second trip was made during the first six months of 1917 when he secured many specimens from Tortuga Island and from the northern and northwestern parts of the Republic of Haiti. In
November of the same year he made a third trip, this time covering southwestern Haiti and Cayemites Island. From February to October, 1919, he visited the Samaria Peninsula once more, and worked to the southwest toward Duverge. In the spring and early summer of 1920, Doctor Abbott visited Gonaives Island and some small villages in the vicinity of Furey, Haiti. The three expeditions taken since that time have all been to the Samana Peninsula, from which district very rich collections have been secured where formerly few specimens had been obtained”. .
Cochran, D. M. (1924). Notes on the Herpetological Collections made by Dr. W. L. W. Abbott on the Island of Haiti. Proceedings of the United States National Museum, 66(2543), 1–15.
Crother, B. I., & Schwartz, A. (Eds.). (1999). Caribbean amphibians and reptiles. Academic Press.
Hedges, S. B., Powell, R., Henderson, R. W., Hanson, S., & Murphy, J. C. (2019). Definition of the Caribbean Islands biogeographic region, with checklist and recommendations for standardized common names of amphibians and reptiles. Caribbean Herpetology, 1–53. https://doi.org/10.31611/ch.67
Henderson, R. W., & Schwartz, A. (1984). A guide to the identification of the amphibians and reptiles of Hispaniola (Vol. 4). Milwaukee Public Museum; /z-wcorg/.
Henderson, R., Noeske-Hallin, T. A., Ottenwalder, J. A., & Schwartz, A. (1987). On the Diet of the Boa Epicrates striatus on Hispaniola, with Notes on E. fordi and E. gracilis. Amphibia-Reptilia, 8, 251–258. https://doi.org/10.1163/156853887X00298
Maclean, W. P., Kellner, R., & Dennis, H. (1977). Island lists of West Indian amphibians and reptiles. Smithsonian Herpetological Information Service, 40, 1–47. https://doi.org/10.5479/si.23317515.40.1
Powell, R. (1993). Comments On The Taxonomic Arrangement Of Some Hispaniolan Amphibians And Reptiles. Herpetological Review, 24(4), 135–137.
Reynolds, R. G., & Henderson, R. W. (2018). Boas of the World (Superfamily Booidae): A Checklist With Systematic, Taxonomic, and Conservation Assessments. Bulletin of the Museum of Comparative Zoology, 162(1), 1–58. https://doi.org/10.3099/MCZ48.1
Reynolds, R. G., Niemiller, M. L., Hedges, S. B., Dornburg, A., Puente-Rolón, A. R., & Revell, L. J. (2013). Molecular phylogeny and historical biogeography of West Indian boid snakes (Chilabothrus). Molecular Phylogenetics and Evolution, 68(3), 461–470. https://doi.org/10.1016/j.ympev.2013.04.029
Reynolds, R. G., Puente-Rolón, A. R., Burgess, J. P., & Baker, B. O. (2018). Rediscovery and a Redescription of the Crooked-Acklins Boa, Chilabothrus schwartzi (Buden, 1975), Comb. Nov. Breviora, 558(1), 1–16. https://doi.org/10.3099/MCZ46.1
Schwartz, A. (1970). A new species of large Diploglossus (Sauria: Anguidae) from Hispaniola. Proc. Biol. Soc. Washington, 82(60), 777–788.
Sheplan, B. R., & Schwartz, A. (1974). Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships. Annals of the Carnegie Museum, 45, 57–143.
Snider, A. T., & Bowler, J. K. (1992). Longevity of reptiles and amphibians in North American collections. Society for the Study of Amphibians and Reptiles.
Welch, K. R. G. (1994). Snakes of the world: a checklist. R & A Research and Information ; KCM Books [distributor].