The nominate species was described and named by Edward Drinker Cope (1840-1897), a paleontologist and zoologist of great stature. He worked at the Smithsonian, the British Museum, Jardin des Plantes (in Paris) and became the Curator of the Philadelphia Academy of Natural Sciences.
Holotype
Academy of Natural Sciences of Philadelphia: Syntypes no. 10237 and no. 10239, New Providence Island, Bahamas, from the collection of Dr. H.C. Wood, Jr.
* Cope, in his 1886 "An Analytical Table of the Genera of Snakes", considered Homalochilus, Pelophilus, Dendrophilus and Piesigaster all synonyms of Chilabothrus.
** Cope also considered Epicrates versicolor a synonym of Chilabothrus strigilatus .
Common Name
Bahamas Boa, Bahamian Boa.
Description and taxonomic notes
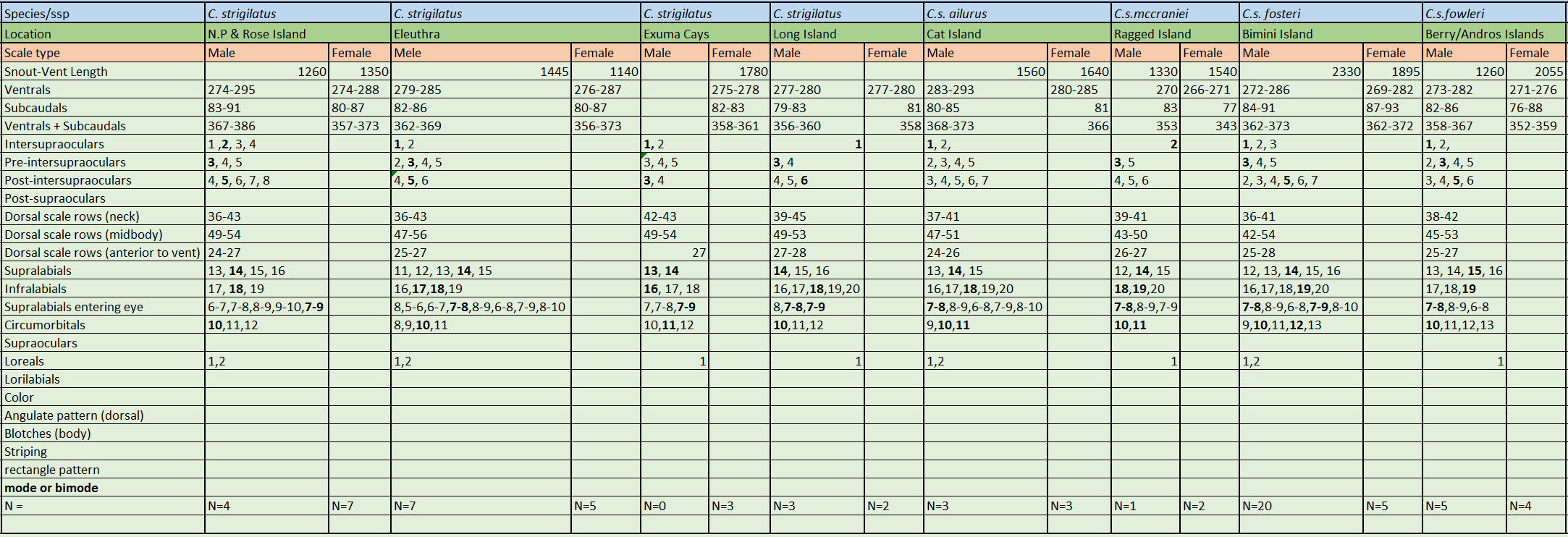
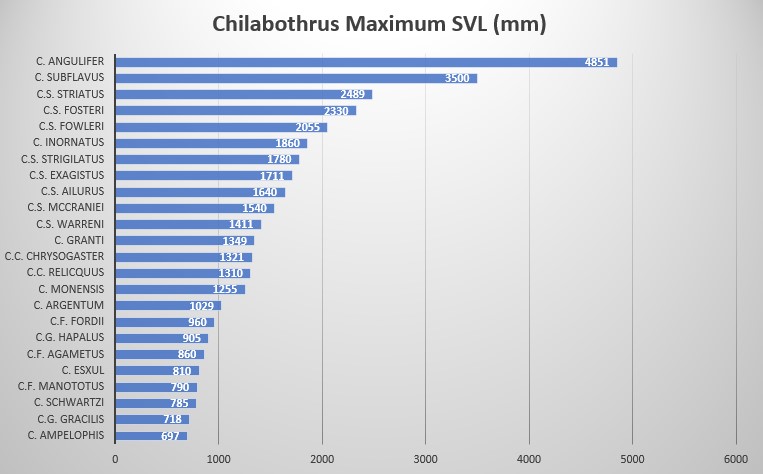
Chilabothrus strigilatus is sexually dimorphic and largely arboreal. The species is characterized by its moderate size, reaching lengths of up to 2330 mm SVL. Males are generally 1445 mm SVL and females 1780 mm SVL. Hanlon reported a 2235 mm long female from New Providence Island . Ventrals in males are 274-295 and 274-288 in females, 1 intersupraocular (modally) [but New Providence specimens are 2 intersupraoculars (modally)], bordered by 3 scales anteriorly (modally) and by 5 scales posteriorly [but Long Island specimens are 6 (modally)], supralabials 14 (modally) with 2 or 3 scales entering the eye (7-8 or 7-9), circumorbital series in 10 scales (modally) and 1 loreal (modally).
Head scale view, C. strigilatus, Eluethera. Photo Sebastian Hoefer
Currently five subspecies are recognized. Cope initially described Chilabothrus strigilatusstrigilatus based on one specimen from New Providence, Bahamas . Chilabothrus strigilatus fosteri was described by Barbour solely on the basis of its coloration . Three of the other subspecies (C. s. ailurus, C. s. mccraniei C. s. fowleri) were described by Sheplan and Schwartz based on a combination of various morphological characters . Their monumental work might be the most significant contribution to our understanding of the morphological characteristics of all Chilabothrus species, known at the time, and the variations of these species within the genus. At the time of their writing, the systematics of the Antillean forms were in a “chaotic state”. They examined living specimens of all Antillean taxa (at the time) except C. chysogaster and C. monensis. Click here to see their publication. We consider it a must read.
A recent molecular phylogeny came to conclude that the two subspecies C. s. fosteri and C. s. fowleri form a single clade. It was therefore suggested to treat C. s. fosteri and C. s. fowleri as a single form termed Chilabothrus strigilatus cf. fosteri . While the authors did a thorough analysis and used only specimens with a confirmed provenance, it should also be noted that the sample size for the study was overall relatively small; the Bimini and Berry Island boas consisted of a total of four specimens (1 Bimini Boa, 3 Berry Island Boas). While this analysis delivered results consistent in itself, we are cautious to follow their argumentation for several reasons and thus continue to treat the currently described subspecies of C. strigilatus as such. The traditional view of the subspecies was recently employed by the lead author of the study in another paper . Thus, until further research provides strong evidence to support the 2013 viewpoint, we consider all subspecies described so far as valid.
Evolution
Subfossil remains from the late Holocene were found by Steadman and co-workers on Eleuthera . Furthermore, fossil remains of a boa larger in size than C. exsul have been found on Abaco. The fossil remains found on Abaco stem from one individual that had lived in between 4200 – 1000 years before present. The authors of the study considered it to be “Epicrates striatus” using the taxonomic status present for the bahamian boas of the strigilatus group at the time of their writing in 2007 .
Researching further they reconsidered their statement and refer to the boa as Chilabothrus cf. exsul. They reason their statement as follows: “the adult fossil vertebrae from Sawmill Sink suggest a midsized Chilabothrus (larger than modern C. exsul, but smaller than C. striatus).” . They however seem to exclude the possibility that this boa might have been a smaller (now extinct) form of C. strigilatus. The finding of this specimen poses the question whether there were several Chilabothrus species present on Abaco at the time, or whether C. exsul underwent a size diminishing process over the course of time. Molecular evidence suggests that Chilabothrus strigilatus is on an isolated evolutionary trajectory since 2.6 MYA .
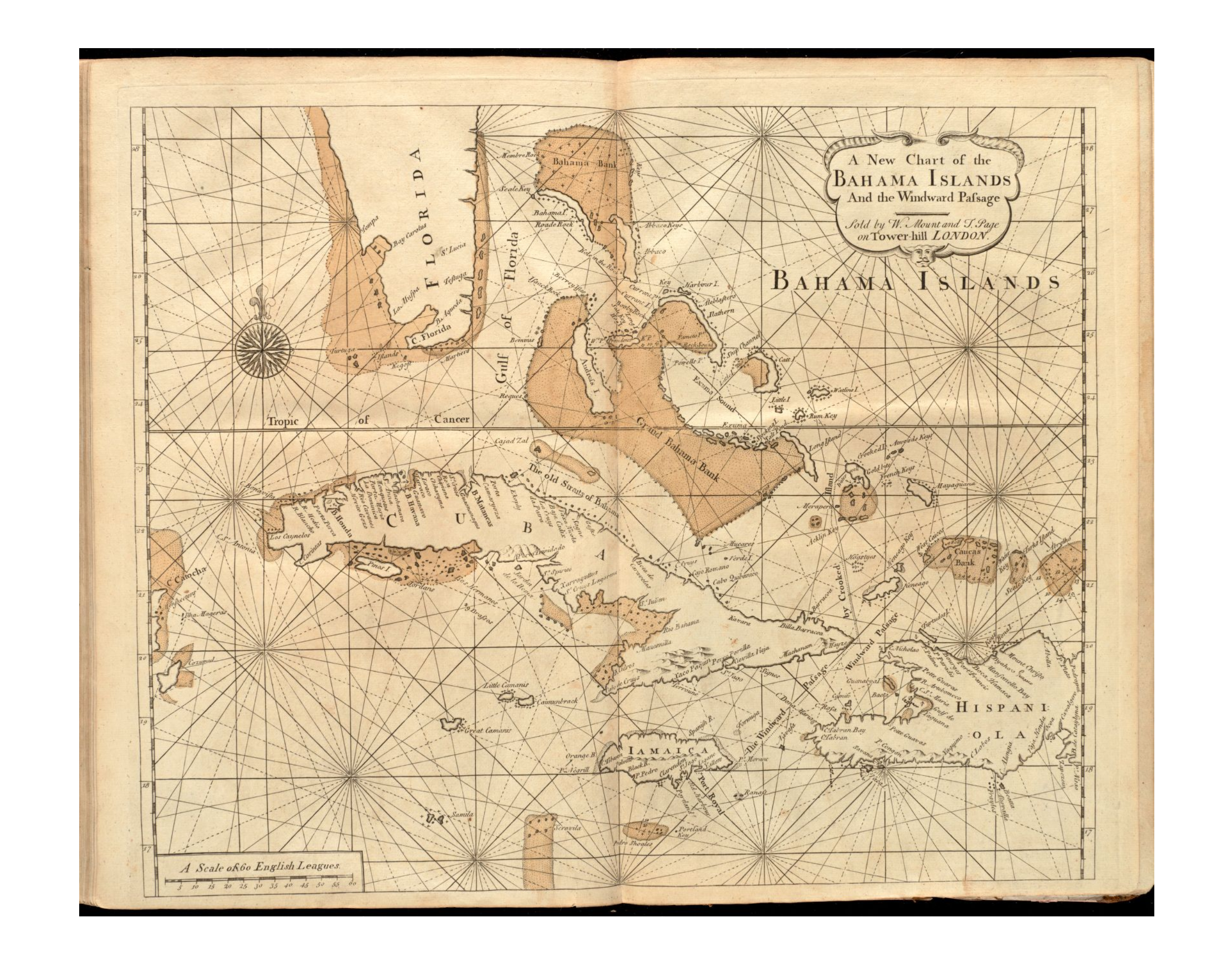
Distribution
All subspecies of Chilabothrus strigilatus are found in the Bahamas. Some occur on a few small islands or cays with a very small surface area. The Bahama Islands consist of Rose Island, New Providence, Long Island, Eleuthra and Exuma Cays.
Chilabothrus s. strigilatus: Bell Island, Compass Cay, Eleuthera Island, Great Exuma Island and Exuma Cays, Hawksbill Cay, Lee Stocking Island, Long Island, New Providence Island, Rose Island.
Juvenile C. s. strigilatus from Long Island. Photo Graham ReynoldsJuvenile Chilabothrus s. strigilatus, Eleuthera. Photo by Sebastian Hoefer
The five subspecies of Chilabothrusstrigilatus occur on different islands of the Bahamas. The flags on the map indicate a type locality of a Chilabothrus strigilatus subspecies. Click on each flag to see which subspecies occurs on that particular island.
Beautiful drawing of Epicrates versicolor from Steindachner, 1863.
** Sheplan and Schwartz remark: “Examination of the detailed scutellar descriptions and figures, as well as the description of the pattern of versicolor, leaves little doubt that this name is a synonym of strigilatus and that the holotype was brought to Europe from some Bahamian island, rather than from Colombia or Hispaniola. If at some future time the Bahamian snakes presently regarded as E. striatus strigilatus are subdivided, then versicolor will require further consideration”. To our knowledge this has not been done.
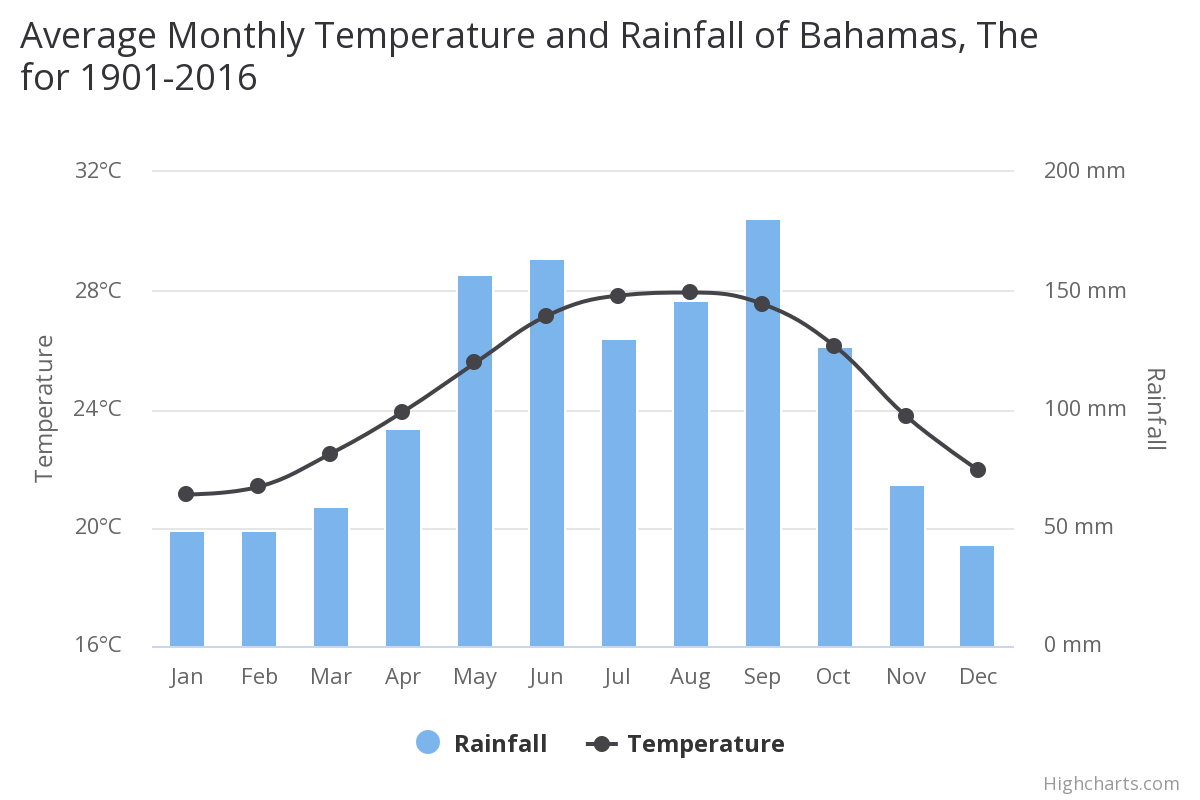
In the Bahamas the mean annual temperature is 24.73°C (76.51° F) and the mean annual precipitation is 1264.46 mm. The islands experience warm, humid conditions year-round, though with more seasonal variations than the Southern Caribbean islands. There are also variations between the islands of the Bahamas, with rainfall falling twice as much in the northwestern islands than in the southeastern islands, and the more northerly islands experiencing temperatures up to 5° cooler than the southern islands.
Average temperatures are fairly high, with the mean daily temperatures fluctuating between 17°C and 32°C (62.6° F and 89.6° F). Mean annual rainfall for the Bahamas varies from about 865 mm to about 1470 mm. Inter-annual variability in climate is strongly influenced by the El Niño Southern Oscillation (ENSO). El Niño episodes bring warmer and drier conditions between June and August. Located in the heart of the Atlantic hurricane belt, the Bahamas is also subject to hurricanes and tropical cyclones especially during the August – November period.
Mean temperatures have increased by around 0.5°C since 1960, at an average rate of 0.11°C per decade. Bahamian data show that the mean daily maximum temperature for July has increased at a rate of 2°C per 100 years, and more recently at a rate of 2.6°C per 100 years. There have been statistically significant increases in the frequency of ‘hot’ days and nights, and decreases in ‘cold’ days and nights during the period 1973-2008.
There is seasonal variation in the rate of temperature increase, with the rate being most rapid in the warmest seasons, June-August and September-November, having rates of 0.13 and 0.15°C per decade respectively. There is also variation between islands, where the rate of warming is more rapid in the northeastern islands compared to the southwestern islands.
C. s. strigilatus is found in hardwood forest, mangrove and in scrub/bush. It is more tolerant of arid conditions than striatus on Hispaniola. As a largely-arboreal boa it can be found in trees and bushes. It has adjusted to the manmade habitat well; so much so it is often removed from garages, out buildings, etc., and released in more natural habitat. (Johnson pers. comm.).
C.s. strigilatus on New Providence. Photo Paul MitzelfeltChilabothrus s. strigilatus on Eleuthera. Photo Sebastian Hoefer
Longevity
Like all members of the species for which we have longevity records, Chilabothrus strigilatus are long lived: 20+ years is the average not the exception. J. Wagner (pers. comm.) has a male C. strigilatus that he acquired as an adult in the 1990’s and assumes this animal is more than 30 years old by now.
Behavior
Chilabothrus strigilatus are very active boas; they are excellent climbers as noted in Henderson (XXX) . A boa was found as high as five meters up in a tree. In Terraria, the boas are always curious observing the area outside their terraria carefully and take the opportunity to explore. They seem to be docile, yet when initially handled they appear a bit nervous, but calm down when finally in the hands of the keeper. They don’t tend to bite but rather flee when disturbed.
Diet
In the wild the young take small lizards (Anolis) as first foods, like the remainder of the genus (with the exception of C. angulifer). With growth the boas eventually shift their diet to rodents (introduced Mus and Rattus), birds, fowl and their eggs. Sheplan and Schwartz assume that the large number of specimens they collected from Hatchet Bay, Eleuthera, is due to the extensive poultry-raising industry at that locality . Hoefer and Colleagues investigated the diest of road killed specimens on Eleuthera across four habitat types. They confirmed the dietary shift and found young snakes exclusively preying on lizards whereas in adults they found mainly mammalian prey .
In captivity the neonates start on ectothermic prey such as Anolis sagrei. It is possible that one or two from a litter might take a live pink mouse but they are the exception to the rule. Once the babies are feeding regularly they can be switched to pm by scenting them with anole skin, blood from the tail of an anole or chick fuzz. At this point the growing boas take appropriately sized rodents (rats and mice), chicks and quail.
Reproductive biology
The species has been kept from the 1960s through the 2000s by various zoos and academic collections. Reproductive data from these has not always been published, however Slavens reports the following reproduction data : C.s. strigilatus: 1985 JACF 0.0.27 born Sept 29/85. 0.0.37 born Oct 9/85 from two different females.
In addition we found the following records. The first breeding of C. s. strigilatus at the Jacksonville Zoo in 1976 and probably the first breeding of C. s. strigilatus at the San Antonio Zoo. They reported to have bred the “Eleuthera Island Boa” producing 30 young in 1969 .
Chilabothrus strigilatus is able to produce large litters of small young. Hanlon reported a litter of 51 young from a female Chilabothrus strigilatus strigilatus of 1676 mm length. The animal was caught on New Providence Island the 19th of June and was placed with several other smaller Boas in a cage. The female refused to eat from the time of its capture until 19th of September .
The same author observed breeding activity in his group of C. s. strigilatus: a female 2235 mm copulated in captivity at least three times with two different males on different days (March 13, 14 and April 8.). The male mated several other females until April 13. The smaller females were subsequently released into the wild. The large female was kept and gave birth on October 17th, resulting in 33 live young, 4 dead and “some 14 of the yolk-like-structures”. The author concludes from his observations a gestation range of 192 to 218 days. Given the length of gestation, we suspect the female was provided with sub-optimal temperatures, thus the extended gestation period.
Trutnau reported a litter of 53 young . More recently (2016) Scott Johnson reported on Facebook a litter of 63 young from his female C. s. strigilatus. To our knowledge, this is the maximum litter sized produced by the species and also by any member of the genus Chilabothrus. The boa originated from New Providence Island and was kept in captivity on the Bahamas until parturition, whereupon it was released back to the wild.
In captivity the boas mate in the months March through May; parturition occurs in the months September through November. Neonate SVL is 318 mm-500 mm .
Gravid Bahamas Boa, C. strigilatus. This boa gave birth on 12 October 2020.
T. Crutchfield, R. Potts, J. Murray, S. Woodward, B. Chambers, M. Saina occasionally produce litters from different lines and subspecies. Those results are listed in the tables below.
C. s. strigilatus (supposed origin New Providence)
C. s. strigilatus (supposed origin New Providence)
C. s. strigilatus (supposed origin New Providence)
J. Murray reports breedings of C.s. strigilatus on:
M. Saina reports breedings of C. s. strigilatus on:
1 October, 2021 (27 offspring, no stillborn, no egg masses)
Litter of Bahamas Boas, C. strigilatus, being born.Female Bahamas Boa eating unfertilized ovum. She also consumed several stillborn babies.Litter of newborn Bahamas Boas, C. strigilatus.
Reproduction is triggered, according to Tolson and Huff, by regularly misting the enclosure during the months March through May.
Captive management
Nursery for neonate Bahamas Boas, C. strigilatus.Baby Bahamas Boa (C. strigilatus) taking an anole scented pink mouse.
Conservation status, threats and population size in nature
The 22 July, 2015 IUCN rating of LC is described as follows:
The species is endemic to the Great Bahama Bank, where it occurs on many islands and cays, and therefore has an extent of occurrence (EOO) well above the threshold for a threatened listing under criterion B1 and occurs in numerous locations and it is not considered to be severely fragmented. The number of mature individuals of the species is inferred to be declining as a result of direct persecution and road mortality throughout most of its range; however, population reductions are not known to approach 30% decline over the last 15 years or anticipated to do so over the coming 15 years (= three generations). Research regarding population trends is considered a high priority for the species, and if these reveal substantial declines, reassessment would become urgent.
While describing C.s. fosteri, Barbour comments on the rarity of C.s. strigilatus on New Providence when previously it was to be found in abundance . Hoefer (2021) found 28 DOR (dead on the road) specimens of C.s. strigilatus on Eleuthera during a ten month period from May, 2019 through May, 2020 Translating to a mortality rate of 2.8 boas per month . In a later study the researchers found that even more boas are victims of roads. They calculated 5.2 road kills per month amounting to an annual loss of 62.4 boas .
The Data from Eleuthera and Chilabothrus s. strigilatus are but the second time road mortality data has been collected on an island for a member of the Genus Chilabothrus; the first being DOR data for C. exsul on Abaco Island, where Giery and colleagues calculated the loss of 1.6 Boas per week (= 83.2 Boas per year) .
A short video of C.s. strigilatus on New Providence can be found here.
At the time of this writing (Sept. 2019) IUCN lists the conservation status of this species as least concern, however, based on the observation that the number of mature individuals is decreasing they conclude that the population is decreasing as a whole. Unfortunately a lack of data makes it impossible to say with certainty how steep the decline is, if any, and what the current population total is. As for all Island fauna, the restricted range they inhabit is a constant threat for species survival.
Data provided by Buckner et al. was used to generate the table below:
Taxon
Number of islands
combined total surface area in km2
Chilabothrus s. strigilatus
9
1548.57
Chilabothrus s. ailurus
2
390
Chilabothrus s. fosteri
4
17.86
Chilabothrus s. fowleri
4
6211.75
Chilabothrus s. mccraniei
2
4.42
The total surface area of the islands as outlined here should not be falsely interpreted as suitable habitat. Detailed analyses about population size and suitable habitats are missing. We list this table to visualize potential threats as we consider it astonishing and worrying at the same time, that some subspecies formed on very small islands. It is worrying that the cane toad Rhinella marina has been recorded from New Providence as well as from Abaco Islands in the West Indies . Considering the devastating impact it had on the Australian herpetofauna, we hope that the West Indian species are less impacted by its occurrence.
Interestingly, the Bahamas Boa Chilabothrusstrigilatus has been recorded outside its natural habitat in Florida . The authors of the study found 2 other Chilabothrus species, C. granti and C. inornatus. The origin of the C. granti and C. inornatus could be determined with certainty, since they were blind passengers in a cargo shipment of electronic waste from Puerto Rico. The authors assume that C. strigilatus originated from the pet trade, since it was discovered in Florida’s Dade County at a facility not involved in international shipping. There is no evidence given, that these alien species could have survived in Florida and form self sustaining populations. There is also no evidence that more animals of these species are present in Florida or elsewhere outside their natural habitat.
The CIA World Factbook lists the following environmental threats for The Bahamas: coral reef decay; solid waste disposal .
The maps below illustrate the extent of habitat destruction and alteration due to development and agriculture.
Rose Island (C.s. strigilatus)Nassau (C.s. strigilatus)New Providence map of 1700. Map Library of CongressEleuthra (C.s. strigilatus)Early map of the Bahamas, 1737.
To add an historic note to the map from 1737 is a passage from Catesby (1743) regarding the Bahamas iguana population and its role in the trade and food chain. He writes, “These Guana’s are a great Part of the Subsistance of the Inhabitants of the Bahama Islands, for which Purpose they visit many of the remote Kays and Islands in their Sloops to catch them, which they do by Dogs trained up for that Purpose, which are so dexterous as not often to kill them, which if they do, they serve only for present spending; if otherwise they sew up their Mouths to prevent their biting, and put them into the Hold of their Sloop till they have catched a sufficient Number, which they either carry alive for Sale to Carolina, or Salt and barrel up for the Use of their Families at Home. These Guana’s feed wholly on Vegetables and Fruit, particularly on a Kind of Fungus, growing at the Roots of Trees, and of this and others of the Anoma Kind.”
Population in captivity
The Bahamas Boa was kept and bred for more than half a century in public and private collections. The International Zoo Yearbook provides lists of institutional breedings. The list for the 1969 breeding records contains a record from San Antonio Zoo. They mention 30 young of the “Eleutera Island Boa” .
Today, at least three distinct lines exist in captivity. Supposedly originating from Eleuthera Island, New Providence Island and an unknown locale, based on the pattern of the adults and all the offspring.
On display in these Zoos
We are unaware that this species is kept in any zoological institution worldwide.
Beautiful Bahamians
Chilabothrus strigilatus strigilatus
Several lines of Bahamas Boa Chilabothrus strigilatus strigilatus are are present in captivity. These slender snakes make perfect display animals in naturalistic terraria.
Chilabothrus strigilatus strigilatus
Chilabothrus strigilatus strigilatus Bahamas Boa, supposedly originating from Eleuthera Island
Chilabothrus strigilatus strigilatus
Chilabothrus strigilatus strigilatus Large female Bahamas Boa
Chilabothrus strigilatus strigilatus
Chilabothrus strigilatus strigilatus Bahamas Boa consuming undeveloped or unfertilized ova after partuition
Chilabothrus strigilatus strigilatus
Bahamas Boa Chilabothrus strigilatus strigilatus The skin of this beautiful male appears shiney / waxy. This is typical for males during breeding season.
Chilabothrus strigilatus strigilatus
Bahamas Boa Chilabothrus strigilatus strigilatus often spent times on branches and are a pleasure to watch
Chilabothrus strigilatus strigilatus
Chilabothrus strigilatus strigilatus Bahamas Boa, supposedly originating from New Providence Island
Chilabothrus strigilatus strigilatus
Chilabothrus strigilatus strigilatus Bahamas Boa, supposedly originating from New Providence Island
Chilabothrus strigilatus strigilatus
This beautiful Bahamas Boa displays a stunning zebra-like pattern and color variation
Chilabothrus strigilatus strigilatus. Photo by Tom Crutchfield
Chilabothrus strigilatus strigilatus. Photo by Tom Crutchfield
Chilabothrus s. strigilatus, Eleuthera, Photo by Sebastian Hoefer
Chilabothrus s. strigilatus, Eleuthera, Photo by Sebastian Hoefer
Chilabothrus s. strigilatus, Eleuthera. Photo by Sebastian Hoefer
Chilabothrus s. strigilatus, Eleuthera. Photo by Sebastian Hoefer
Chilabothrus s. strigilatus, Eleuthera. Photo by Sebastian Hoefer
Chilabothrus s. strigilatus, Eleuthera, Photo by Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Chilabothrus s. strigilatus on Eleuthera, Photo Sebastian Hoefer
Prev
Next
Several Pictures of Chilabothrus strigilatus from various sources (link outs)
Barbour, T. (1937). Third list of Antillean reptiles and amphibians. Bulletin of the Museum of Comparative Zoology at Harvard College., 82, 77–166. https://www.biodiversitylibrary.org/part/14803
Barbour, T. (1935). A second list of Antillean reptiles and amphibians. Zoologica : Scientific Contributions of the New York Zoological Society., 19(3), 77–141. https://www.biodiversitylibrary.org/part/203717
Barbour, T. (1904). Batrachia and Reptilia from the Bahamas. Bulletin of the Museum of Comparative Zoology at Harvard College., 46, 55–61. https://www.biodiversitylibrary.org/part/13038
Barbour, T. (1941). A new boa from the Bahamas. New England Zoological Club, Proceedings., 18, 61–65. /z-wcorg/.
Boulenger, G. A. (1893). Catalogue of the snakes in the British Museum (Natural History). Printed by order of the Trustees; /z-wcorg/.
Buckner, S. D., Franz, R., & Reynolds, R. G. (2012). BAHAMA ISLANDS AND TURKS & CAICOS ISLANDS. In Island lists of West Indian amphibians and reptiles (Vol. 2, pp. 93–110).
Cope, E. D. (1862). Synopsis of the species of Holcosus and Ameiva, with diagnoses of new W. Indian and S. American Colubridae. Proceedings of the Academy of Natural Sciences of Philadelphia, XIII, 60–150. https://doi.org/10.1093/nq/s5-VI.146.318i
Cope, E. D. (Edward D. (1887). List of the Batrachia and Reptilia of the Bahama Islands. Proceedings of the United States National Museum., 10(645), 436–439. https://doi.org/10.5479/si.00963801.10-645.436
Cope, E. D. (1886). An Analytical Table of the Genera of Snakes. Proceedings of the American Philosophical Society, 23(124), 479–499.
Currie, D., Wunderle, J. M., Freid, E., Ewert, D. N., & Lodge, D. J. (2019). The natural history of the Bahamas: a field guide. Comstock Publishing Associates, an imprint of Cornell University Press.
Garman, S. (1887). On West Indian reptiles in the Museum of Comparative Zoology at Cambridge, Mass. Proceedings of the American Philosophical Society, 24(126), 278–286. /z-wcorg/.
Hanlon, R. W. (1964). Reproductive Activity of the Bahaman Boa (Epicrates striatus). Herpetologica, 20(2), 143–144. http://www.jstor.org/stable/3890592
Hedges, S. B., Powell, R., Henderson, R. W., Hanson, S., & Murphy, J. C. (2019). Definition of the Caribbean Islands biogeographic region, with checklist and recommendations for standardized common names of amphibians and reptiles. Caribbean Herpetology, 1–53. https://doi.org/10.31611/ch.67
Hoefer, S., Mills, S., Pinou, T., & Robinson, N. J. (2021). What the Dead Tell Us about the Living: Using Roadkill to Analyze the Diet and Endoparasite Prevalence in Two Bahamian Snakes. Ichthyology & Herpetology, 109(3). https://doi.org/10.1643/h2020141
Hoefer, S., Robinson, N. J., & Pinou, T. (2021). Size matters: Sexual dimorphism in the pelvic spurs of the Bahamian Boa (Chilabothrus strigilatus strigilatus). Herpetology Notes, 14, 201–203.
Krysko, K. A., Somma, L. A., Smith, D. C., Gillette, C. R., Cueva, D., Wasilewski, J. A., Enge, K. M., Johnson, S. A., Campbell, T. S., Edwards, J. R., Rochford, M. R., Tompkins, R., Fobb, J. L., Mullin, S., Lechowicz, C. J., Hazelton, D., & Warren, A. (2016). New Verified Nonindigenous Amphibians and Reptiles in Florida through 2015, with a Summary of over 152 Years of Introductions. IRCF Reptiles & Amphibians, 23(2), 110–143.
Maclean, W. P., Kellner, R., & Dennis, H. (1977). Island lists of West Indian amphibians and reptiles. Smithsonian Herpetological Information Service, 40, 1–47. https://doi.org/10.5479/si.23317515.40.1
Mertens, Robert. (1939). Herpetologische Ergebnisse einer Reise nach der Insel Hispaniola, Westindien. Vittorio Klostermann; /z-wcorg/.
Reynolds, R. G., & Henderson, R. W. (2018). Boas of the World (Superfamily Booidae): A Checklist With Systematic, Taxonomic, and Conservation Assessments. Bulletin of the Museum of Comparative Zoology, 162(1), 1–58. https://doi.org/10.3099/MCZ48.1
Reynolds, R. G., Niemiller, M. L., Hedges, S. B., Dornburg, A., Puente-Rolón, A. R., & Revell, L. J. (2013). Molecular phylogeny and historical biogeography of West Indian boid snakes (Chilabothrus). Molecular Phylogenetics and Evolution, 68(3), 461–470. https://doi.org/10.1016/j.ympev.2013.04.029
Reynolds, R. G., Collar, D. C., Pasachnik, S. A., Niemiller, M. L., Puente-Rolón, A. R., & Revell, L. J. (2016). Ecological specialization and morphological diversification in Greater Antillean boas. Evolution, 70(8), 1882–1895. https://doi.org/10.1111/evo.12987
Reynolds, R. G., Giery, S. T., Jesse, W. A. M., & Quach, Q. N. (2016). Preliminary Assessment of Road Mortality in Chilabothrus exsul, the Northern Bahamas Boa. Caribbean Naturalist, 34.
Reynolds, R. G., Puente-Rolón, A. R., Burgess, J. P., & Baker, B. O. (2018). Rediscovery and a Redescription of the Crooked-Acklins Boa, Chilabothrus schwartzi (Buden, 1975), Comb. Nov. Breviora, 558(1), 1–16. https://doi.org/10.3099/MCZ46.1
Rosen, N. (1911). Contributions to the fauna of the Bahamas. Lunds Univ. Arsskr., 7, 1–72.
Schwartz, A., & Henderson, R. W. (1985). A guide to the identification of the amphibians and reptiles of the West Indies exclusive of Hispaniola. Milwaukee Public Museum.
Sheplan, B. R., & Schwartz, A. (1974). Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships. Annals of the Carnegie Museum, 45, 57–143.
Steadman, D. W., Albury, N. A., Kakuk, B., Mead, J. I., Soto-Centeno, J. A., Singleton, H. M., & Franklin, J. (2015). Vertebrate community on an ice-age Caribbean island. Proceedings of the National Academy of Sciences, 112(44), E5963–E5971. https://doi.org/10.1073/pnas.1516490112
Steadman, D. W., Albury, N. A., Mead, J. I., Soto-Centeno, J. A., & Franklin, J. (2018). Holocene vertebrates from a dry cave on Eleuthera Island, Commonwealth of The Bahamas. The Holocene, 28(5), 806–813. https://doi.org/10.1177/0959683617744270
Steadman, D. W., Franz, R., Morgan, G. S., Albury, N. A., Kakuk, B., Broad, K., Franz, S. E., Tinker, K., Pateman, M. P., Lott, T. A., Jarzen, D. M., & Dilcher, D. L. (2007). Exceptionally well preserved late Quaternary plant and vertebrate fossils from a blue hole on Abaco, The Bahamas. Proceedings of the National Academy of Sciences, 104(50), 19897–19902. https://doi.org/10.1073/pnas.0709572104
Steindachner, F. (1863). Über eine neue Epicrates-Art aus Columbien. Denkschriften Der Kaiserlichen Akademie Der Wissenschaften / Mathematisch-Naturwissenschaftliche Classe Wien, 22, 10.
Stejneger, L. (1905). Batrachians and Land Reptiles of the Bahama Islands. In G. B. Shattuck (Ed.), The Bahama Islands (The Johns Hopkins Press, pp. 329–337). The Macmillan Company.
Stimson, A. F. (1969). Boidae (Boinae + Bolyeriinae + Loxoceminae + Pythoninae). De Gruyter; /z-wcorg/.
Stull, O. G. (1935). A Checklist of the Family Boidae. Proceedings of the Boston Society of Natural History, 40, 387–408.
Trutnau, L. (1988). Schlangen im Terrarium: Haltung, Pflege und Zucht; in 2 Bänden. 1: Ungiftige Schlangen (3., durchges. u. erw. Aufl). Ulmer.
Walls, J. G. (1999). The Living Boas: a complete guide to the boas of the world. T.F.H. ; Kingdom; /z-wcorg/.
Welch, K. R. G. (1994). Snakes of the world: a checklist. R & A Research and Information ; KCM Books [distributor].